-
【听课笔记】复旦大学遗传学_04连锁与交换
课程地址:复旦大学遗传学
四、连锁与交换
4.1 连锁交换
4.1.1 连锁与交换的概念
-
连锁 (
linkage):同一染色体上的基因以及它们所控制的性状结合在一起传递的现象。 -
交换 (
crossing over):减数分裂中,同源染色体的非姐妹染色单体在染色体某处形成交叉 (chiasma) 并发生局部互换,导致互换位置两侧的连锁基因发生重组的现象。 -
连锁与交换定律 (
law of linkage and crossing-over):位于同一条染色体上的两个或两个以上的基因倾向于联合在一起进行传递 , 当这些基因之间的某处位置发生了交换,就会产生重组类型的配子。(遗传第三大定律) -
位于同一条染色体上的两个基因在减数分裂中始终联系在一起同时传递给配子,形成两种亲本型配子,比例为 1:1 的现象,称为完全连锁。

-
位于同一条染色体上的两个基因在减数分裂中或者联系在一起同时传递给配子,形成亲本型配子;或者发生非姐妹染色单体间的交换后重新组合,形成两种重组型配子;且重组型配子比例小于亲本型配子比例,称为不完全连锁。

-
交换发生在减数分裂中
同源染色体配对发生在偶线期,同源染色体交换发生在粗线期,在双线期能看到交叉的形成,所以,遗传物质的交换发生在交叉之前,交叉只是同源染色体发生交换的位置留下的痕迹,交叉是交换的结果,即遗传学上的交换发生在细胞学上的交叉出现之前。
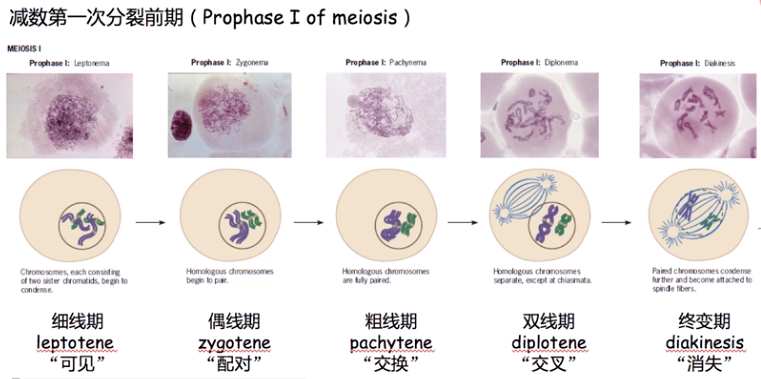
-
联会复合体
在粗线期中出现联会复合体 ( senaptonemal complex ) 介导同源染色体的配对和非姐妹染色单体的交换。交换发生在交叉 (chiasma) 处,交叉在粗线期形成,在染色体上不断向末端移动以完成染色体的交换,到浓缩期结束才消失。

4.1.2 连锁现象的发现
● 香豌豆两对相对性状杂交试验
- 1906 年英国学者 Bateson 和 Pannett 利用香豌豆的两对相对性状(花色、花粉粒形状)进行了两种杂交组合实验
花色︰紫花 § 对红花 § 为显性
花粉粒形状︰长花粉粒 (L) 对圆花粉粒 I 为显性
杂交组合 1∶紫花 长花粉粒 × 红花 圆花粉粒;

杂交组合 2∶紫花 圆花粉粒 × 红花 长花粉粒。 - 结果:F2 与亲本组合相同的类型多,重新组合的类型偏少,原为同一亲本的两个性状在子二代中常有联系在一起的现象,这说明来自同一亲本的基因有较多的在一起传递的可能。
● 连锁交换定律的发现
- 1912 年摩尔根利用果蝇的两对相对性状进行正反交实验
体色∶灰体 (vg+) 对黑体 (vg) 是显性
翅形︰长翅 (b+) 对残翅 (b) 是显性

- 正反交结果不同,后来发现这两种性状位于同一染色体上,正交时由于 F1 雄蝇的基因完全连锁,所以没有重组型个体出现;反交时,F1 雌蝇中两个非等位基因之间发生了交换(不完全连锁),所以 F2 中有少量重组型。
● 重组率与遗传距离
- 在判定两个基因之间是否连锁 时,我们将 F1 代双杂合子进行测交,如果后代有四种基因型 (1:1:1:1) 则是自由组合;如果后代中仅有两种亲本基因型或者重组型比率低于亲本型则说明两个基因之间相互连锁。
- 两基因之间连锁的程度可以用测交子代中的重组率 (
recombination frequency, RF) 进行估计︰
重组率 = 重组型数目 /(亲本型数目+重组型数目)
△ 将 1% 的重组值记为一个单位,称一个厘摩,即 1cM,这个距离也被称为遗传距离。
△ 由于交换和重组发生的可能性与基因之间的距离有密切关系,因此,遗传距离对了解染色体上的基因真实的物理距离有参考意义。

4.1.3 三点测交
- 三点测交 (
three point test cross) ,也称为三点试验,是由摩尔根建立的计算 3 个基因之间的重组值/连锁关系的简便方法。即利用三杂合体与三隐形纯合个体测交,在同一次杂交实验中就能获得三个基因之间的重组值关系。 - 1 个三点测交相当于 3 个“两点测交”。但相比两点测交,还可以计算双交换值,并且降低遗传背景和环境条件的干扰。
● 三点测交案例
-
X 染色体上的三个隐性突变基因∶sn3(卷刚毛); w(白眼);m(小翅)
-
sn3 w m / + + + 雌果蝇和 sn3 w m/ Y 雄果蝇杂交
-
雌蝇在减数分裂中发生重组与交换,产生 8 种雌配子,而雄蝇的 X 染色体和 Y 染色体不能配对交换,只有 2 种配子:一种是 X 染色体上带有三个隐性基因的配子,一种是带有 Y 染色体的配子。所以 F1 雌果蝇产生的配子的表型与 F2 个体的表型一致,通过 F2 个体的表型可推算 F2 雌果蝇产生的 8 种配子的类型和数目。

-
F2 雌果蝇产生的 8 种配子的类型和及其统计结果
左图:F2 雌果蝇产生的 8 种配子的类型和数目
右图:每种配子的类型,及非等位基因之间是否存在交换(-未交换,+交换)

- 计算三个基因间的重组率,就可以画出三个基因在染色体上的连锁图。

- 在连锁的两个非等位基因之间会发生多次交换,两个基因之间距离越长,发生交换的次数越多。重组的配子只能检测到奇数交换,而检测不到双交换,由于双交换比例最低,因此数目最少的两种配子为双交换类型。双交换是两个单交换同时发生的结果,因此,双交换值也可以根据两个单交换的乘积计算。

- 校正值∶被低估的双交换频率。
(4+4)×2 / 1000 = 1.6%
33.5%+1.6%= 35.1% - 并发率(并发系数)= 双交换率的观察值 / 单交换率观察值的乘积
0.8% / ( 15.1% × 20% ) = 0.26 - 干涉 = 1 - 并发率 = 1-0.26 =0.74
- 在三点测交中,中间的基因与两侧的基因同时分开的机会很小,如果两个基因对间的单交换并不影响邻近两个基因对间的交换,预期的双交换的频率应等于两个单交换频率的乘积,干涉率为 0。干涉率为 1 时,表示干涉是完全的,意味着两个基因之间没有双交换发生。
- 干涉 反映的是染色体上一次单交换对于邻近发生的另一次单交换的影响程度。
★ 连锁群和连锁图
- 如果 A 与 B 连锁,C 与 B 连锁,则 A 与 C 连锁。
- 如果 A 与 B 连锁,B 与 C 不连锁,则 A 与 C 不连锁。
- 连锁群 (
linkage group) : 位于一对同源染色体上的具有一定连锁关系的基因群。连锁群数目等于单倍体染色体数 (n)。 - 连锁图 (
linkage map) : 又称为遗传学图,是根据染色体上的基因之间的相互交换值和排列顺序制定的、表明连锁基因的位置和相对距离的线性图谱。 - 利用重组值绘制的遗传图谱,会出现根据重组值得到的距离小于遗传距离的现象。遗传距离大于真实重组值的原因:在距离较长的基因之间发生了多次交换,产生非重组型配子。
4.2 真菌的连锁分析
4.2.1 四分子分析

-
粗糙链孢霉 (Neurospora crassa) 属于真菌类中的子囊菌,是一种良好的遗传分析材料。
-
四分子分析的优势:
单倍体世代:基因型直接在表现型中表现出来;
二倍体世代:同源染色体之间具有配对、交换现象,能进行重组分析;
减数分裂产生的四个子囊孢子呈直线顺序排列。 -
对链孢霉子囊孢子进行基因分析的方法称为四分子分析。
-
四分子分析可以很好地判断减数分裂中有无交换、 干涉,确定发生交换的染色单体。
-
链孢霉的生活史:

-
利用四分子分析进行单基因分析时,可以将着丝粒看做一个基因座位。
4.2.2 着丝粒作图
- 着丝粒作图 (
centromere mapping) : 计算某一基因与着丝粒之间的遗传距离,并据此在染色体上进行基因定位的作图方法。(将着丝粒看作一个基因座位,计算基因与着丝粒之间的重组关系和遗传距离。)
● 案例:
-
野生型的子囊孢子是黑色的,但赖氨酸缺陷型的孢子成熟较慢,是灰色的。故而可以利用颜色表型可以判断等位基因型。
-
野生型 (原养型
lys+) 和赖氨酸缺陷型 (营养缺陷型,lys-) 的杂交,分析杂合子形成的子囊中子囊孢子的排列情况。 -
观察发现,lys+lys- 杂合子的子囊中会出现 6 种不同的子囊孢子排列,包括 2 种非交换类型和 4 种交换类型:


-
非交换型子囊的形成方式:第一次分裂分离型

-
交换型子囊的形成方式:第二次分裂分离型


-
基因与着丝粒遗传距离的计算
在第一次分裂分离的子囊中基因位点与着丝点之间没有发生交换。
在第二次分裂分离的子囊中,四条染色单体中有两条发生交换。
用 N1、N2 分别表示第一次、第二次分裂分离的子囊数。
发生交换的染色单体数等于第二次分裂分离子囊数的 2 倍,即 2N2。
所有染色单体的总数是子囊数的 4 倍,即 (4N1 + 4N2)。
则,基因与着丝粒遗传距离 = 发生交换的染色单体数 / 所有染色单体的总数

5 /(2×5 + 2×9) = 0.18
4.2.3 两个基因的连锁作图
- 两对基因在染色体上的位置关系可能是:
(1) 无连锁,位于两条不同染色体上
(2) 有连锁,位于同一条染色体着丝粒两端
(3) 有连锁,位于同一条染色体着丝粒的一侧 - 一对基因有 6 种子囊类型,那两对基因具有 6×6 = 36 种子囊类型,根据不同类型子囊的数目可以判定基因的连锁关系和遗传距离。
● 案例:
- nic 突变型(烟酸依赖型)和 ade 突变型 ( 腺嘌呤依赖型)
- nic + 与 + ade 杂交,计 1000 个子囊,基因型和数量。
- 两对基因杂交虽然有 36 种子囊类型,排除减数分裂过程中子囊孢子的随机取向等因素后,可将 36 种子囊类型归纳为 7 种子囊型,3 种子囊类别:PD(亲二型,与亲本相同)、NPD(非亲二型,与亲本不相同的重组类型)、T(四型,包括两种亲本型和 2 种重组型)

- 分别计算单个基因与着丝粒的距离

- 判断两个基因的关系

■ 第一种类型:两基因位于不同染色体上,且两基因都和各自的着丝粒紧密连锁,基因和着丝粒之间不发生交换。子囊类型只有 PD、NPD 两种,且理论比例为 1:1。而上述结果 PD 有 898 个,远大于 NPD,因此不属于此种类型。

■ 第二种类型:如果两基因位于不同染色体上,且基因和各自染色体着丝粒之间发生交换。产生 PD、NPD 和 T 型 3 种子囊类型,且 PD 比例低于 50%。而实验结果 PD 有 898 个,远大于 50%,因此不属于此种类型,说明这两个基因不是自由组合的,存在连锁关系。

■ 第三种类型:如果两基因位于同一染色体上,且两基因之间完全连锁,无交换。只产生 PD 子囊类型,没有 NPD 和 T 型子囊。而实验结果 NPD 和 T 型子囊都存在,因此排除此种类型。

■ 第四种类型:如果两基因位于同一染色体上,且两基因之间不完全连锁,发生一次交换。产生 PD 子囊类型,比例大于 50%,出现 T ,但无 NPD,因此排除此种类型。

■ 第五种类型:如果两基因位于同一染色体上,且两基因之间不完全连锁,除发生一次交换外,还有涉及到多条染色单体的双交换。产生 PD 子囊类型,比例大于 50%,出现 T、NPD,符合此种类型。

- 可根据各基因与着丝粒之间的交换型和非交换型子囊数,计算基因与着丝粒的距离,但需要对双交换进行校正。根据以上分析可以做出以下判断:
 ■ 亲二型 = 80.8% > 50%,排除两基因自由组合的可能。
■ 亲二型 = 80.8% > 50%,排除两基因自由组合的可能。
■ 出现 T,排除完全连锁,有交换;又出现 NPD, 有双交换。
因此,两基因连锁,并位于着丝点一侧。
■ 分别计算 nic 和 ade 与着丝粒的距离,分别为 5.05、9.30
■ 计算 nic 与 ade 的距离为 5.20
■ 按照基因在染色体上呈直线排列的原则,ade 与着丝粒的距离应该为:5.05 + 5.20 = 10.25 > 9.30,说明在计算 ade 与着丝粒的距离时,双交换被掩盖了,也说明这一实验中,配子的形成涉及多条染色单体的双交换。

■ 由于两个基因间的双交换,导致根据重组值计算的距离与遗传距离不符的结果,可以通过补足双交换值来校正。之前计算 nic 和 ade 与着丝粒的距离时是将所有第二次分裂分离的子囊相加除 2 倍总子囊数,这样遗漏了少量发生过双交换的子囊(2、4、6、7),据此计算双交换值为 0.95%,0.95+9.30=10.25,即完全符合基因的直线排列原理。


- 根据上述分析,作 nac 和 ade 之间的遗传连锁图。

4.3 遗传标记与遗传图谱
- 1%交换值 = 1cM ≈ 1000kb
- 人基因组全长 3300cm,如果两个标记之间的距离为 1cM,则需要 3300 个标记;如两个标记之间的距离为 2-5cM,则需要 660-1650 个标记。
- 绘制遗传连锁图的方法很多,但能使用的遗传标记很少,最初使用的标记多数是根据表型性状确定的。发现 DNA 序列可以作为遗传标记后,根据连锁标记建立的连锁群和绘制的遗传连锁图越来越清晰的展现在人们面前。
4.3.1 遗传标记
- 遗传标记 (
Genetic marker) 用于连锁分析、基因定位、遗传作图等的可遗传的生物标记;目前发现的可识别的生物标记包括可识别的个体形态、细胞特征、免疫性状或者特定的 DNA 序列等。 - 经典遗传学常选用形态标记和细胞学标记,它们是早期遗传学研究中容易分辨,能够在上下世代之间传递的特征标记。这些标记不能满足绘制精密连锁图的需求。
- 现代遗传学则多选用生化和分子水平标记,它们在染色体上的覆盖度更高,多态性好,也易于检测。
- 三种分子遗传标记:
■ 第一代–RFLPRestriction Fragment Length Polymorphism 限制性片段长度多态性 (1987 年,H.Donis-Keller , 人基因组 RFLP 连锁图)
■ 第二代–SSLPSimple sequence length polymorphism 简单序列长度多态性(1992 年,Welssenbanch , 人基因组 STR 连锁图)
■ 第三代–SNPSingle nucleotide polymorphism 单核苷酸多态(1996 年,Lander , 人基因组 SNP 连锁图)
限制性片段长度多态性 RFLPs
- 原理:通过检测 DNA 在限制性内切酶酶切后的特定 DNA 片段大小来判断基因的多态性。基因突变(点突变、缺失和插入等)可导致基因序列中原有限制酶切位点的消失或新酶切位点的出现,从而引起不同 DNA 在同一限制酶切割时,产生不同长度的 DNA 片段。

- 人基因组中大约含有 10 万个 RFLPs, 是广泛使用的第一代遗传标记。RFLP 通常使用 Southern 印迹方法进行检测。
- 一个家系不同成员 RFLP 标记的结果

- RFLPs 的特点
■ 优点
(1)无表型效应,不受环境条件和发育阶段的影响;
(2)在等位基因之间是共显性的,不受杂交方式的影响;
(3)非等位的 RFLP 标记之间互不干扰;
(4)源于基因组 DNA 的自身变异,在数量上不受限制。
■ 缺点
(1)依赖基因序列限制性酶切位点的变异,只能检测到一种杂合的两种形式,即有或无,且每个位点杂合子在人群中的频率通常不会超过 50%,当被测个体为纯合时,利用 RFLP 标记无法得到需要的多态信息;此外,整个基因组中,这类标记数目有限,分布不均匀,因此,遗传多态性水平相应比较低。
(2)由于需要序列信息进行 PCR 分析,另外还需要相应的探针,因此对于那些缺乏相应分子信息的物种来说应用这种标记是很困难的。
简单序列长度多态性 SSLPs
-
基因组中的串联重复序列核心序列(重复单元)的重复次数在人群中存在变异,形成了序列长度的一种多态现象,是广泛应用的第二代等位标记。
-
人类基因组中最常见的 SSLP 包括:
■ 小卫星 DNA:重复单位6-25bp, 也称为数目变异串联重复VNTRvariable number of tandem repeat,主要存在于染色体靠近端粒处,在不同个体之间存在串联数目的变异。
■ 微卫星 DNA:重复单位2-6bp, 也称为 短串联重复STR, simple tandem repeat。同一类微卫星 DNA 可分布于基因组不同位置,由于重复次数不同、重复程度的不完全造成每个位点的多态性。
-
SSLP 通常使用 Southern 印迹方法进行检测。
-
DNA 指纹 (
DNA Fingerprints)
■ 1984 年 Jefferys 等人利用分离得到的人小卫星 DNA 制作探针,与核基因组的酶切片断杂交,得到多条带组成的杂交图谱。
■ 选用的探针是肌红蛋白基因第一个内含子中的串联重复序列,重复单位长度为 33bp,从人的基因文库中筛选出 8 个克隆,序列分析后发现,每个克隆都含有一个长 0.2 ~ 2.0kb,重复单位重复 3 ~ 29 次组成的小卫星 DNA。尽管重复单位的长度和序列不相同,但都含有一段相同的 16bp 核心序列,以这个 16bp 序列为一个重复单元,选用重复 29 次的小卫星序列做探针(33.15),用这个探针与人基因组酶切片段进行杂交,在低严谨条件下,杂交产生由 10 多个条带组成的杂交图谱。用另一个小卫星探针(33.6)进行测试,也能获得类似的图谱。这些图谱在不同个体之间完全不同,就像指纹一样因人而异,被称之为 DNA 指纹,又名遗传指纹图谱。DNA 指纹反应了小卫星 DNA 的多态性。产生 DNA 指纹的过程,叫做 DNA 指纹分析。

-
DNA 指纹的特点
■ 多位点性: 一方面,小卫星 DNA 的串联数目存在差异,酶切片段或 PCR 产物大小有多态性。另一方面,小卫星 DNA 内部含有相同或相似的核心序列,能够被同一基因探针识别,容易检测。
■ 高变异性:两个无血缘关系的个体具有相同 DNA 指纹图谱的概率极低。
■ 稳定遗传:DNA 指纹条带的遗传遵循孟德尔遗传规律。
单核苷酸多态性 SNP

-
在基因组水平上由单个核苷酸的变异所引起的 DNA 序列多态性,称作单核苷酸多态性
SNP。 -
定义一个 SNP 要求任何一种核苷酸在群体中的频率不小于 1%,否则可视为突变。
-
尽管人类的基因密码大部分是相同的,但不同人之间仍然在每 1000 个核苷酸中就有一个 SNP , 这是人与人之间遗传差异的重要组成部分。
-
SNP 与 RFLP 和 SSLP 的区别
■ 与 RFLP 和 SSLP 等标记相比,SNP 不再以“长度”的差异作为检测手段,而是直接以序列的差异作为标记。
■ 但 SNP 单个基因座的多态性和 RFLP 一样,只有二态,但多个 SNPs 构成的多态可以弥补 SNPs 二态性的缺陷。
■ 二态,即一对同源染色体上同一位点的 2 个碱基对发生变异的状态,在人群中可以有 3 种基因型。
如上图例子所示,这 3 种基因型包括:两条染色体均是 T-A 碱基对的纯合子、均是 G-C 碱基对的纯合子、TA/GC 杂合子。从理论上看,每个 SNP 位点都可以有 4 种不同的变异形式,但实际上经常发生的只有 2 种(转换或颠换),大多数情况下,C 都变成 T,变成 A/G 的概率很小,所以一般认为 SNP 是二态性的。
■ 和微卫星相比,微卫星可以表现出大于二态性的多态性,但是 SNP 在基因组上分布密集,掩盖了它二态性的缺点,其优点远远超过微卫星,成为目前最理想的第三代分子标记。 -
SNP 的优点
■ SNP 数量大,分布广;
■ 以寡核苷酸杂交为基础的 DNA 芯片技术易于实现自动化批量检测,可用计算机分析结果。
■ 由于 SNPs 是人群差异的主要来源之一,基于全基因组的 SNP 扫描对复杂疾病的标记基因或易感基因筛选具有重要意义。
4.3.2 基因定位
-
基因定位包括将基因定位于某一特异的染色体上,以及测定基因在染色体上线性排列的顺序和距离。
-
在基因位置不详,但基因型可知的情况下,可以利用分子遗传标记和家系材料对基因进行定位。
-
家系连锁分析法
■ 以两代或两代以上的家系材料为基础,观察标记位点与致病基因位点在家系内是否呈“共分离”, 并利用优势对数计分法 (Lod, log odds score) 计算出遗传距离和连锁程度。
■ Lod = lg 两基因座连锁且相距 θ 值的似然性 两基因座不连锁但相距 θ 值的似然性 \frac {两基因座连锁且相距 θ 值的似然性}{两基因座不连锁但相距 θ 值的似然性} 两基因座不连锁但相距θ值的似然性两基因座连锁且相距θ值的似然性
■ 在判断基因与某个标记之间是否连锁时,Lod 值将在连锁情况下观察到实验数据的似然性与纯属偶然观察到相同数据的似然性进行比较。 -
案例:Huntington disease 的基因定位
■ 亨廷顿舞蹈病:常染色体显性遗传;外显率高;容易获得大的家系;临床表现为痴呆和舞蹈样动作,发病人群多为中年人,患者最终变得不能说话、走路、吞咽,直至死亡。病理:部分脑基底神经节处有明显的神经元缺失。
■ 定位方案:收集家系资料,开发全基因组 RFLP 探针 (200 多个),逐个分析,获得与致病基因连锁的 RFLP,获得基因位置的信息。


-
-
相关阅读:
GSMA SGP.01协议学习
修改 MySQL 最大连接数
Vector 模拟实现
k8s docker 中部署think php 并搭建php websocket
django理解03 数据库引入
MySQL的select语句
worthington是谁?worthington抑制剂对酶活性的影响研究
小龙虾优化算法COA求解不闭合SD-MTSP,可以修改旅行商个数及起点(提供MATLAB代码)
为什么禁止MyBatis批量插入几千条数据使用foreach?
AD域安全攻防实践(附攻防矩阵图)
- 原文地址:https://blog.csdn.net/zea408497299/article/details/126037911